Hormonal HealthSystematic Review
Perimenopause Nutrition: Alcohol, Fibre, Protein, and Omega-3 -- A Functional Medicine Framework
Evidence:Systematic reviews
Chris MassambaFunctional Nutritionist & Health Coach · Dip CNM · FMCHC30 May 2026Updated 9 June 202634 min readMedical disclaimer
Key points
- Perimenopause is a 4-to-10-year hormonal transition that affects approximately 13 million women in the United Kingdom at any given time, yet nutritional guidance during this window remains inconsistent and often overlooked in conventional primary care.
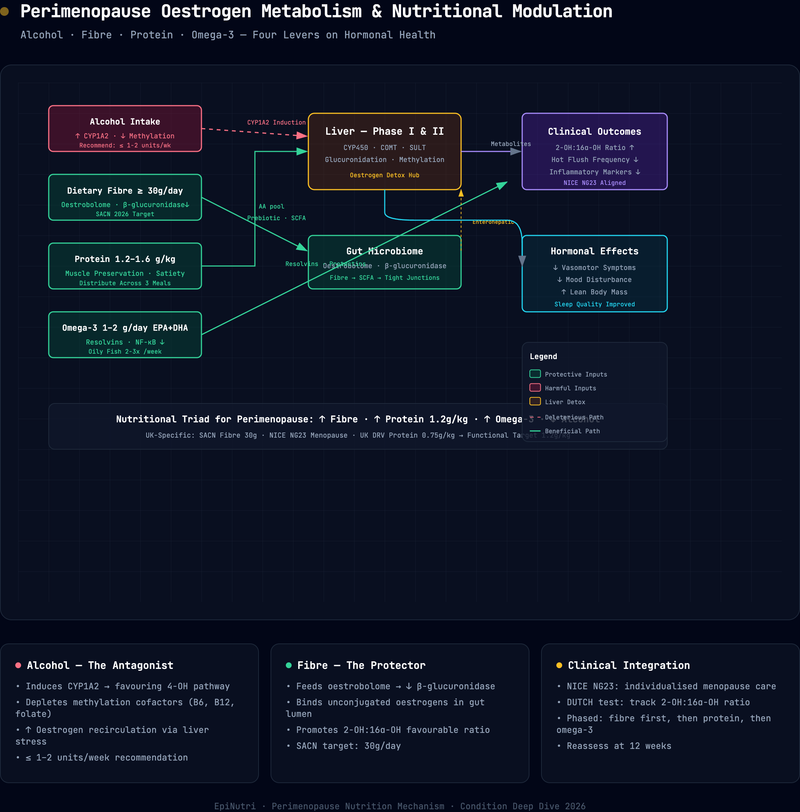
- Four dietary levers -- alcohol reduction, fibre optimisation, adequate protein, and omega-3 fatty acid intake -- address the root-cause mechanisms of the most common perimenopausal complaints: vasomotor symptoms, body composition change, mood instability, and systemic inflammation.
- Alcohol, even at moderate intake, elevates circulating oestrogen and its genotoxic metabolites, increasing breast cancer risk by approximately 10 per cent for every 10 grams of ethanol consumed daily.
- The gut oestrobolome -- the collection of bacterial genes capable of metabolising oestrogen -- is directly modulated by dietary fibre. Meeting the SACN recommendation of 30 grams per day supports balanced oestrogen recirculation and excretion.
- Protein requirements during perimenopause rise above the standard UK Reference Nutrient Intake (0.75 g/kg/day) to at least 1.2 g/kg/day, and up to 1.6 g/kg/day for physically active women, to counter accelerated sarcopenia driven by declining oestrogen.
- Omega-3 fatty acids (EPA and DHA) exert anti-inflammatory effects that may attenuate the low-grade systemic inflammation underpinning cardiovascular risk escalation, joint pain, and mood disturbance during the menopausal transition.
- NICE guideline NG23 (2015, updated 2024) provides the clinical framework for identification and management; this article complements -- not replaces -- those recommendations with a functional medicine nutritional lens.

The core question: what can nutrition do for perimenopause?
Perimenopause is a physiological transition, not a disease. But its symptoms -- vasomotor instability, accelerated muscle loss, mood disruption, systemic inflammation -- are profoundly influenced by what a woman eats and drinks during these years. The evidence points to four dietary levers that address the underlying physiology rather than simply masking symptoms: alcohol, fibre, protein, and omega-3 fatty acids.
NICE NG23 provides the clinical framework for pharmacological management, and HRT can be transformative. But the guideline offers comparatively limited nutritional guidance. It does not discuss oestrogen metabolism through the oestrobolome, does not quantify protein requirements for sarcopenia prevention, and does not address the specific impact of alcohol on oestrogen recirculation. This article fills that gap.
Sarah (not her real name), a 47-year-old solicitor from Manchester, arrived at her appointment carrying a folded sheet of paper. On it she had written five words: "I do not feel like me."
Over the previous eighteen months, her periods had become irregular -- sometimes arriving every twenty-one days, sometimes skipping six weeks entirely. She woke at three in the morning drenched in sweat, her heart racing, three or four nights a week. She had gained a stone around her midsection despite running three times a week and eating what she called "a healthy diet." Her joints ached. Her patience, once her professional asset, had frayed. She described crying in the car park after routine meetings.
Her GP had tested her thyroid and full blood count -- both within range -- and offered a low-dose SSRI for mood. Her blood glucose was 5.8 mmol/L, nudging toward the prediabetic threshold. Nobody had asked about her diet in any detail. Nobody had explained the hormonal mechanics of what was happening. Nobody had connected the wine she drank most evenings -- "just two glasses, everyone does" -- to the very symptoms disrupting her life.
Sarah's story is not unusual. Approximately three-quarters of women going through the menopausal transition experience symptoms, roughly a quarter describe them as severe, and evidence suggests that a third endure them for years beyond their final period.
Clinical pearl: When a perimenopausal patient says "I do not feel like me," take that statement seriously. It is not vague -- it reflects the cumulative impact of hormonal volatility on mood, cognition, body composition, and sleep. The four dietary levers in this article address the upstream physiology behind that experience.

Perimenopause in the United Kingdom
Definition and staging
Perimenopause is the transitional phase preceding menopause, defined by the Stages of Reproductive Aging Workshop (STRAW+10) as the period during which menstrual cycle variability increases and reproductive hormone levels fluctuate erratically (Harlow et al., 2012). The STRAW+10 system divides this transition into an early stage (cycle length variation of seven or more days) and a late stage (episodes of amenorrhoea lasting sixty or more days). Menopause itself is diagnosed retrospectively after twelve consecutive months of amenorrhoea.
Epidemiology
The average age of menopause in the United Kingdom is 51, with perimenopause typically beginning between 45 and 55 years of age. Current estimates suggest that approximately 13 million women in the UK are perimenopausal or postmenopausal at any given time -- roughly one-third of the entire female population. Women aged 45 to 55 are the fastest-growing demographic in the UK workforce, making perimenopause not merely a clinical concern but an economic and occupational one.
Approximately one in a hundred women experiences premature ovarian insufficiency (menopause before the age of 40), and these individuals face additional cardiovascular, skeletal, and cognitive risks that warrant earlier and more intensive intervention.
Risk factors for problematic perimenopause
Not every woman experiences disabling symptoms, but several factors increase likelihood and severity: smoking (advances menopause by approximately two years), higher visceral adiposity, alcohol consumption, sedentary behaviour, chronic stress (HPA-axis dysregulation may compound hormonal volatility), and poor dietary quality -- particularly ultra-processed food intake, low fibre, and inadequate protein.

Pathophysiology: oestrogen decline and metabolic shifts
The hormonal cascade
The menopausal transition is driven by the progressive depletion of ovarian follicles. As the follicular pool diminishes, the ovaries produce less inhibin B, reducing the negative feedback on the anterior pituitary. Follicle-stimulating hormone (FSH) rises, initially driving compensatory increases in oestradiol. This creates the characteristic hormonal volatility of early perimenopause: wild fluctuations in oestradiol -- sometimes supraphysiological, sometimes profoundly low -- against a backdrop of rising FSH and declining progesterone (Santoro et al., 2021).
As the transition advances, oestradiol production falls more consistently, and the ratio of oestrone (the weaker postmenopausal oestrogen, produced primarily in adipose tissue) to oestradiol shifts. This is not a simple decline; it is a reorganisation of the entire endocrine landscape, with downstream effects on virtually every organ system.
Metabolic consequences
The metabolic consequences of declining oestrogen are substantial (Santoro et al., 2021; Erdelyi et al., 2023):
- Insulin sensitivity decreases: oestrogen enhances insulin signalling in skeletal muscle and hepatocytes. Its withdrawal promotes insulin resistance, visceral fat accumulation, and an atherogenic lipid profile. Continuous glucose monitoring can make these metabolic shifts visible in real time.
- Lean mass declines: oestrogen has anabolic effects on skeletal muscle. Its decline accelerates sarcopenia. Women may lose approximately 0.5-1.0 per cent of muscle mass per year during the transition, with rates increasing postmenopausally (Erdelyi et al., 2023).
- Systemic inflammation increases: oestrogen suppresses pro-inflammatory cytokines (IL-6, TNF-alpha). Its decline unmasks low-grade chronic inflammation -- "inflammageing" -- underpinning cardiovascular disease, osteoarthritis, and mood disturbance.
- Bone resorption accelerates: oestrogen inhibits osteoclast-mediated bone resorption. The perimenopausal years see the most rapid bone mineral density loss.
At a glance -- Metabolic consequences of oestrogen decline:
Insulin resistance increases. Lean mass declines (0.5-1.0% per year). Systemic inflammation unmasks. Bone resorption accelerates. Gut microbiome composition shifts. Each of these is addressable through the four dietary levers discussed below.
- Gut microbiome composition shifts: the oestrogen-gut axis is bidirectional. Declining oestrogen alters the gut microbiome, and an altered microbiome changes oestrogen metabolism. This feedback loop, mediated by the oestrobolome, is central to the fibre discussion below (Baker, Al-Nakkash and Herbst-Kralovetz, 2017). For patients whose oestrobolome needs more than dietary fibre alone, the 5R gut restoration framework provides a structured approach.
The Communication node in the Functional Health Matrix
Within the EPINUTRI Functional Health Matrix, perimenopause maps primarily to the Communication node, which encompasses hormonal signalling, receptor sensitivity, and the intercellular messaging systems that coordinate metabolic and reproductive function. A declining Communication score during the perimenopausal years reflects the progressive disruption of oestrogen, progesterone, and their downstream signalling cascades. Nutritional interventions that modulate oestrogen metabolism, reduce inflammation, and support neurotransmitter synthesis directly address this node.
Figure: Functional Health Matrix -- Perimenopausal Pattern
Description: A radar chart displaying the seven Functional Health Matrix nodes. The Communication node is scored 1-2 (highlighted in amber as the primary therapeutic target), reflecting disrupted hormonal signalling. Defence scores 2-3 (reflecting the unmasking of systemic inflammation as oestrogen declines). Structural Integrity scores 2-3 (reflecting accelerated bone resorption and sarcopenia). Assimilation may score 2-3 if oestrobolome dysbiosis is present. Energy, Transport, and Biotransformation typically score in the 3-4 range unless additional comorbidities are present.
Legend: Each node scored 1-5 (1 = severely compromised, 5 = optimal). Total out of 35. Nodes scoring 1-2 are primary therapeutic targets. The Communication node is the index node for perimenopause; Defence and Structural Integrity are commonly affected secondary nodes.
Interpretation: The perimenopausal FHM pattern characteristically shows a suppressed Communication node driving secondary effects in Defence (inflammation) and Structural Integrity (bone, muscle). This distinguishes it from primary inflammatory conditions (low Defence, preserved Communication) or metabolic presentations (low Energy, variable Communication). Each of the four dietary levers in this article maps to a specific node: alcohol reduction and fibre support Communication; protein supports Structural Integrity; omega-3 supports Defence.
Conventional versus functional medicine approach
The conventional pathway
The conventional medical approach to perimenopause, as outlined in NICE NG23 (2015, updated 2024), is pragmatic and evidence-based within its scope. It recommends:
- Diagnosing perimenopause clinically in women aged 45 and over based on symptoms and menstrual change, without routine laboratory testing.
- Offering hormone replacement therapy (HRT) as the primary pharmacological intervention for vasomotor symptoms, after individualised risk-benefit discussion.
- Considering cognitive behavioural therapy (CBT) for mood and vasomotor symptoms.
- Addressing cardiovascular risk factors and bone health.
This approach is appropriate and valuable. HRT, when indicated, can be transformative. However, NICE NG23 offers comparatively limited guidance on nutrition beyond general healthy-eating advice. The guideline does not discuss oestrogen metabolism through the oestrobolome, does not quantify protein requirements for sarcopenia prevention, and does not address the specific impact of alcohol on oestrogen recirculation.
The functional medicine lens
Functional medicine does not oppose HRT or any evidence-based pharmacological intervention. Rather, it asks a different question: what are the modifiable dietary and lifestyle factors that influence the underlying physiology of the transition, regardless of whether HRT is also used?
This approach identifies four primary nutritional levers for perimenopause, each targeting a specific pathophysiological mechanism:
- Alcohol: modulates oestrogen metabolism and recirculation, affects sleep architecture, and increases breast cancer risk.
- Fibre: shapes the gut oestrobolome, supports oestrogen excretion, and improves metabolic markers.
- Protein: counters oestrogen-driven sarcopenia, supports thermogenesis, and preserves functional capacity.
- Omega-3 fatty acids: attenuate systemic inflammation, support cardiovascular health, and may improve mood.
These are not fringe interventions. They are evidence-based dietary modifications that address root causes rather than downstream symptoms. The remainder of this article examines each lever in detail.
Root cause analysis
1. Alcohol: the hidden oestrogen driver
The mechanism
Alcohol affects oestrogen metabolism through multiple pathways. Ethanol increases aromatase activity, the enzyme that converts androgens to oestrogens in peripheral tissues, particularly adipose tissue. It impairs hepatic oestrogen clearance, prolonging the half-life of circulating oestradiol. And it shifts oestrogen metabolite ratios toward the genotoxic 4-hydroxyoestrone pathway, which generates quinone intermediates capable of forming depurinating DNA adducts (Playdon et al., 2018).
In the Women's Health Initiative Observational Study (N = 1,864), alcohol consumption was positively associated with circulating parent oestrogen concentrations in postmenopausal women. The association was dose-dependent and independent of body mass index, suggesting a direct metabolic effect rather than an adiposity-mediated one (Playdon et al., 2018).
From the literature: "Moderate alcohol consumption during adult life, including as few as three to six drinks per week, was associated with a small but statistically significant increase in breast cancer risk." -- Chen et al., JAMA 2011
The risk data
The dose-response relationship between alcohol and breast cancer is one of the most robust findings in cancer epidemiology. In the Nurses' Health Study (N = 105,986 women, 28-year follow-up), Chen et al. (2011) reported that as few as three to six alcoholic drinks per week were associated with a statistically significant increase in breast cancer risk. Each additional 10 grams of ethanol per day -- roughly one small glass of wine or half a pint of standard-strength beer -- increased risk by approximately 10 per cent.
More recently, Sohi et al. (2024) conducted a systematic review and meta-analysis of prospective cohort studies, confirming the dose-response relationship and finding no evidence of a safe threshold for alcohol consumption with respect to breast cancer risk. The risk was consistent across beverage types: wine conferred no protection relative to beer or spirits.
For perimenopausal women, this evidence has particular urgency. The fluctuating and sometimes supraphysiological oestrogen levels of the transition create a hormonal environment in which additional exogenous oestrogen-elevating exposures -- including alcohol -- may amplify cumulative risk. Moreover, alcohol disrupts sleep architecture, fragments slow-wave sleep, and may worsen the nocturnal vasomotor symptoms that already impair restorative sleep during perimenopause (Santoro et al., 2021).
Clinical recommendation
The evidence does not support "moderate drinking" as a benign lifestyle choice during perimenopause. Whilst a blanket prohibition is unlikely to be adhered to and may damage the therapeutic relationship, practitioners should:
- Quantify current intake honestly (many women underestimate consumption).
- Communicate the 10-per-cent-per-10-grams dose-response clearly and without moralising.
- Set a realistic reduction target: ideally fewer than 7 units per week, with at least three alcohol-free days.
- Suggest replacement rituals (sparkling water with lime, kombucha, alcohol-free botanical spirits) to address the habit loop.
- Frame alcohol reduction as a positive oestrogen-management strategy, not a deprivation.
Practice tip: Frame the conversation around oestrogen management, not morality. "Alcohol increases the oestrogen your body is already struggling to regulate -- reducing it is one of the most effective things you can do for your symptoms" lands far better than "you should drink less."
Safety note: If a patient discloses alcohol dependence or consumption above 35 units per week, this is beyond the scope of nutritional counselling. Refer to the GP for assessment and, where appropriate, specialist alcohol services. Abrupt cessation in dependent drinkers can cause dangerous withdrawal.
2. Fibre and the oestrobolome: feeding the hormonal thermostat
The oestrobolome concept
The term "oestrobolome" was introduced by Plottel and Blaser (2011) to describe the aggregate of enteric bacterial genes whose products are capable of metabolising oestrogens. The key enzyme is beta-glucuronidase, produced by specific gut bacteria, which deconjugates oestrogen metabolites in the intestinal lumen. Conjugated (inactive) oestrogens, excreted via bile into the gut, are either eliminated in faeces or reactivated by bacterial beta-glucuronidase and reabsorbed into systemic circulation.
The clinical implication is profound: the gut microbiome acts as a hormonal thermostat. A diverse, well-nourished oestrobolome produces appropriate levels of beta-glucuronidase, maintaining oestrogen homeostasis. Dysbiosis -- driven by low-fibre diets, antibiotic exposure, or chronic stress -- can either reduce beta-glucuronidase activity (leading to excessive oestrogen excretion and relative oestrogen deficiency) or increase it pathologically (leading to excessive oestrogen recirculation and oestrogen dominance) (Baker, Al-Nakkash and Herbst-Kralovetz, 2017).
During perimenopause, when endogenous oestrogen production is already erratic, a dysfunctional oestrobolome compounds the hormonal chaos. Supporting microbial diversity through dietary fibre becomes a direct intervention in oestrogen metabolism, not merely a digestive health measure. For a deeper look at the role of microbial reinoculation in gut restoration, the 5R protocol update on microbiome reinoculation covers the current evidence.
From the literature: "The oestrobolome -- the aggregate of enteric bacterial genes whose products are capable of metabolising oestrogens -- modulates the enterohepatic circulation of oestrogens and affects both circulating and excreted oestrogen levels." -- Plottel and Blaser, Cell Host and Microbe 2011
UK fibre intake: the gap
The Scientific Advisory Committee on Nutrition (SACN, 2015) set the dietary reference value for adult fibre intake at 30 grams per day (measured by the AOAC method). The previous recommendation had been 18 grams per day (by the Southgate method). Average UK intake currently sits at approximately 19 grams per day -- a 37-per-cent shortfall from the target.
For perimenopausal women, this gap has hormonal consequences. Insufficient fibre reduces microbial diversity, impairs short-chain fatty acid production (which has anti-inflammatory and insulin-sensitising effects), and disrupts the oestrobolome's capacity to modulate oestrogen recirculation (Baker, Al-Nakkash and Herbst-Kralovetz, 2017). It also impairs bowel transit time, prolonging the window for oestrogen reabsorption from the colon.
Fibre types and food sources
Not all fibre is equivalent in its effects on the oestrobolome:
- Soluble fermentable fibre (prebiotic): oats, barley, legumes, onions, garlic, leeks, asparagus, bananas. These feed beneficial Bifidobacteria and Lactobacilli, supporting microbial diversity and appropriate beta-glucuronidase activity.
- Insoluble fibre: wholegrains, vegetables, nuts, seeds. These increase faecal bulk and reduce colonic transit time, limiting oestrogen reabsorption.
- Polyphenol-rich fibre sources: berries, flaxseed, cruciferous vegetables. Polyphenols act as selective prebiotics and also support phase II hepatic oestrogen detoxification (glucuronidation, sulphation, methylation).
Flaxseed deserves specific mention. Ground flaxseed provides both soluble fibre and lignans, which are phytoestrogenic compounds that can weakly bind oestrogen receptors and modulate total oestrogenic activity (Baker, Al-Nakkash and Herbst-Kralovetz, 2017). Two tablespoons of ground flaxseed daily (approximately 4 grams of fibre) is a practical addition to the perimenopausal diet.
Clinical recommendation
- Aim for a minimum of 30 grams of fibre per day (SACN target), building gradually over 2-4 weeks to avoid gastrointestinal discomfort.
- Prioritise diversity: at least 30 different plant foods per week (the "30-plant challenge") to maximise microbial diversity.
- Include at least one cruciferous vegetable daily (broccoli, cauliflower, kale, Brussels sprouts, cabbage) for sulforaphane-mediated support of oestrogen detoxification pathways.
- Add 2 tablespoons of ground flaxseed daily for lignans and soluble fibre.
- Increase water intake alongside fibre to support bowel function.
3. Protein: defending lean mass against oestrogen decline
The sarcopenia problem
Oestrogen has direct anabolic effects on skeletal muscle, mediated through oestrogen receptor alpha (ER-alpha) on muscle fibres. As oestrogen declines during perimenopause, protein synthesis rates fall, muscle satellite cell activation decreases, and the balance between muscle protein synthesis and breakdown shifts toward net catabolism (Erdelyi et al., 2023).
This is not an abstract concern. Sarcopenia -- the age-related loss of muscle mass, strength, and function -- accelerates during the menopausal transition. Women may lose approximately 0.5-1.0 per cent of muscle mass per year during perimenopause and early postmenopause (Erdelyi et al., 2023), with corresponding losses in strength that compound fall risk, metabolic rate, insulin sensitivity, and functional independence.
UK reference values versus clinical need
The UK Reference Nutrient Intake (RNI) for protein in adults is 0.75 g/kg body weight per day, as set by the Department of Health's 1991 Dietary Reference Values report. This value was established to prevent deficiency in healthy adults and has not been updated for decades. It does not account for the accelerated muscle protein turnover of the menopausal transition.
The PROT-AGE Study Group position paper (Bauer et al., 2013), whilst focused on adults aged 65 and over, established that older adults require at least 1.0-1.2 g/kg/day to maintain muscle mass, with higher intakes (1.2-1.5 g/kg/day) recommended for those with acute or chronic disease. Nunes et al. (2022), in a systematic review and meta-analysis published in the Journal of Cachexia, Sarcopenia and Muscle, confirmed that protein intakes above 1.2 g/kg/day, combined with resistance exercise, consistently improved lean body mass outcomes in healthy adults.
For perimenopausal women, the clinical consensus in functional medicine and sports nutrition is converging on 1.2-1.6 g/kg/day as the appropriate range, distributed across at least three meals to optimise muscle protein synthesis rates. For broader context on how these protein and fibre targets sit within the evolving dietary guidance landscape for 2025-2030, the alignment article provides the policy background.
From the literature: "Protein intakes above 1.2 g/kg/day, combined with resistance exercise, consistently improved lean body mass outcomes in healthy adults." -- Nunes et al., Journal of Cachexia, Sarcopenia and Muscle 2022
Caution: The UK RNI of 0.75 g/kg/day was set to prevent deficiency in healthy adults and has not been updated for decades. It does not account for the accelerated muscle protein turnover of the menopausal transition. Using this figure as a clinical target for perimenopausal women is likely to be insufficient.
Practical protein targets
For a 70 kg woman, the range translates to:
Practical protein targets
| Target | Daily protein (grams) | Per meal (3 meals) |
|---|---|---|
| UK RNI (0.75 g/kg) | 53 g | 18 g |
| Minimum perimenopausal (1.2 g/kg) | 84 g | 28 g |
| Active/exercising (1.6 g/kg) | 112 g | 37 g |
Legend: Targets are calculated for a 70 kg woman. "UK RNI" is the statutory Reference Nutrient Intake (set 1991, unchanged). "Minimum perimenopausal" reflects the PROT-AGE and functional-medicine consensus. "Active/exercising" applies to women undertaking regular resistance training. "Per meal" assumes three even meals; four smaller meals are also acceptable.
Interpretation: The gap between the UK RNI (53 g/day) and the minimum perimenopausal target (84 g/day) is 31 grams -- equivalent to adding one extra protein-rich meal component per day. Most women eating a standard UK diet are closer to the RNI than the perimenopausal minimum, making this the single most under-addressed nutritional lever in clinical practice.
Reaching 28-37 grams of protein per meal requires deliberate planning. A typical UK breakfast of toast and jam provides 4-6 grams of protein. Upgrading to eggs on wholegrain toast with smoked salmon provides 25-30 grams -- a fivefold increase from a single meal swap.
Protein quality and leucine threshold
Not all protein sources are equivalent for muscle protein synthesis. The leucine threshold -- the minimum leucine dose required to maximally stimulate mTOR-mediated protein synthesis -- is approximately 2.5-3.0 grams per meal. Animal sources (eggs, fish, poultry, dairy) generally provide higher leucine concentrations per gram of protein than plant sources, though combinations of legumes, tofu, and wholegrains can achieve the threshold.
High-leucine UK food sources:
- 150 g chicken breast: 3.6 g leucine, 46 g protein
- 200 g Greek yoghurt: 2.4 g leucine, 20 g protein
- 2 large eggs: 1.5 g leucine, 14 g protein
- 150 g tinned sardines: 2.8 g leucine, 30 g protein
- 200 g cooked lentils: 1.4 g leucine, 18 g protein
Clinical recommendation
- Target a minimum of 1.2 g/kg/day, rising to 1.6 g/kg/day for women engaged in regular resistance training.
- Distribute protein across three to four meals, aiming for at least 25-30 grams per meal.
- Prioritise leucine-rich sources at each meal to cross the anabolic threshold.
- Combine protein targets with progressive resistance training at least twice weekly -- protein without mechanical loading is insufficient to reverse sarcopenia.
- For women who struggle with whole-food protein volume, a high-quality whey or pea protein supplement (20-25 g serving) can bridge the gap, particularly at breakfast.
4. Omega-3 fatty acids: dampening the inflammatory fire
The inflammation mechanism
The withdrawal of oestrogen's anti-inflammatory effects during perimenopause unmasks a state of chronic low-grade inflammation (Santoro et al., 2021). Pro-inflammatory cytokines (IL-6, TNF-alpha, CRP) rise, contributing to:
- Vasomotor symptoms (inflammatory mediators in the thermoregulatory centre).
- Cardiovascular risk escalation (endothelial dysfunction, atherogenic plaque instability).
- Joint pain and stiffness (synovial inflammation).
- Mood disturbance (neuroinflammation affecting serotonin and dopamine signalling).
- Visceral fat accumulation (inflammatory adipokines perpetuating insulin resistance).
Long-chain omega-3 polyunsaturated fatty acids (n-3 PUFAs) -- specifically eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) -- counter this inflammatory cascade through multiple mechanisms. EPA and DHA are incorporated into cell membrane phospholipids, where they competitively displace arachidonic acid (an omega-6 fatty acid) from the substrate pool for cyclooxygenase and lipoxygenase enzymes. This shifts eicosanoid production from pro-inflammatory prostaglandins (PGE2) and leukotrienes (LTB4) toward less inflammatory or actively resolving mediators, including resolvins, protectins, and maresins (Calder, 2017).
From the literature: "Omega-3 fatty acids and inflammatory processes are linked from molecules to man: EPA and DHA compete with arachidonic acid for enzymatic conversion, shifting the balance from pro-inflammatory to pro-resolving mediators." -- Calder, Biochemical Society Transactions 2017
Evidence for perimenopausal benefit
The evidence base for omega-3 supplementation in perimenopause and menopause is growing but heterogeneous:
- Lipid profile: Chen et al. (2023), in a systematic review and dose-response meta-analysis of randomised controlled trials in Clinical Therapeutics, found that omega-3 supplementation significantly reduced triglycerides and may improve HDL-C in postmenopausal women, with dose-dependent effects.
- Vasomotor symptoms: Iqbal et al. (2023), in a systematic review of randomised controlled trials published in Nutrients, found that the evidence for omega-3 supplementation reducing vasomotor symptoms was inconclusive overall, though individual trials showed benefit and no harm was identified.
- Mood and cognition: EPA-predominant omega-3 formulations have shown benefit for depressive symptoms in multiple meta-analyses, though perimenopausal-specific data remain limited.
- Cardiovascular risk: the anti-inflammatory and triglyceride-lowering effects of EPA and DHA are well established and directly relevant to the cardiovascular risk escalation that accompanies oestrogen decline.
UK intake and SACN position
There is no formal UK RNI for EPA and DHA. SACN recommends that the population consume at least two portions of fish per week, one of which should be oily (providing approximately 450 mg of long-chain omega-3 per day). Many perimenopausal women fall below this target, particularly those who do not regularly eat oily fish.
Oily fish sources available in the UK:
- Salmon (farmed Atlantic): 2.2 g EPA+DHA per 140 g portion
- Mackerel: 2.6 g EPA+DHA per 140 g portion
- Sardines: 1.8 g EPA+DHA per 140 g portion
- Herring/kippers: 1.7 g EPA+DHA per 140 g portion
- Anchovies: 1.4 g EPA+DHA per 140 g portion
For women who do not eat fish, algal-derived omega-3 supplements (providing EPA and DHA from microalgae) offer a plant-based alternative.
Clinical recommendation
- Consume at least two portions (280 g total) of oily fish per week, targeting 1-2 g of combined EPA+DHA daily.
- For women who do not eat fish, consider an algal omega-3 supplement providing at least 500 mg combined EPA+DHA daily.
- If supplementing, prioritise EPA-predominant formulations for anti-inflammatory and mood benefits.
- Simultaneously reduce omega-6-dominant seed oils (sunflower, corn, soya) to improve the omega-6:omega-3 ratio and reduce substrate availability for pro-inflammatory eicosanoid production.
- Ensure adequate vitamin D status (SACN recommends 10 micrograms/day for all UK adults), as vitamin D and omega-3 fatty acids have synergistic anti-inflammatory effects. For body-weight-adjusted vitamin D dosing guidance, the late-spring reset article provides the framework.
Treatment algorithm: dietary priorities by symptom cluster
The following framework organises the four nutritional levers by primary symptom presentation, allowing practitioners to prioritise interventions based on the individual patient's most disabling complaints.
Treatment algorithm: dietary priorities by symptom cluster
| Symptom cluster | Primary lever | Secondary levers | Adjunct |
|---|---|---|---|
| Vasomotor (hot flushes, night sweats) | Alcohol reduction | Omega-3, Fibre | Phytoestrogen-rich foods (soya, flaxseed) |
| Body composition (central weight gain, muscle loss) | Protein (1.2-1.6 g/kg/day + resistance training) | Fibre, Alcohol reduction | Time-restricted eating (12-14 h fast) |
| Mood disturbance (anxiety, irritability, low mood) | Omega-3 (EPA-predominant) | Protein (tryptophan for serotonin), Alcohol reduction | Magnesium glycinate, B-vitamin complex |
| Cardiovascular risk (dyslipidaemia, rising BP) | Omega-3 | Soluble fibre, Alcohol reduction | Mediterranean dietary pattern |
| Gut symptoms (bloating, constipation, IBS flare) | Fibre (gradual increase to 30 g/day) | Omega-3, Protein (glutamine-rich sources) | Probiotic foods (kefir, sauerkraut, live yoghurt) |
Legend: "Primary lever" is the intervention with the strongest mechanistic rationale for the given symptom cluster. "Secondary levers" provide additive benefit. "Adjunct" lists complementary strategies with emerging or supportive evidence. Symptom clusters are not mutually exclusive -- most perimenopausal patients present with two or more.
Interpretation: This table allows practitioners to prioritise interventions by the patient's most disabling symptoms. Most patients benefit from all four levers simultaneously, but starting with the primary lever for the dominant symptom cluster improves early adherence and builds confidence for the remaining changes.
Evidence strength by nutritional lever
Evidence strength by nutritional lever
| Intervention | Evidence tier | Key sources |
|---|---|---|
| Alcohol-breast cancer dose-response (10% per 10 g/day) | Strong (meta-analysis of prospective cohorts) | Chen et al. 2011; Sohi et al. 2024 |
| Alcohol elevation of circulating oestrogen | Strong (observational, N = 1,864) | Playdon et al. 2018 |
| Dietary fibre 30 g/day for cardiometabolic benefit | Strong (SACN systematic review) | SACN 2015 |
| Protein 1.2 g/kg/day for lean mass preservation | Strong (systematic review + meta-analysis) | Nunes et al. 2022; Bauer et al. 2013 |
| Omega-3 for triglyceride reduction in postmenopausal women | Strong (dose-response meta-analysis of RCTs) | Chen et al. 2023 |
| Oestrobolome modulation via dietary fibre | Moderate (mechanistic + observational) | Plottel and Blaser 2011; Baker et al. 2017 |
| Omega-3 for vasomotor symptom reduction | Emerging (inconclusive RCTs) | Iqbal et al. 2023 |
| Ground flaxseed for oestrogen modulation | Emerging (small trials + mechanistic) | Baker et al. 2017 |
Legend: Evidence tiers: Strong = supported by meta-analyses of prospective cohorts, systematic reviews, or large RCTs; Moderate = supported by mechanistic data plus observational studies; Emerging = supported by inconclusive RCTs, small trials, or mechanistic evidence only.
Interpretation: The strongest evidence supports alcohol reduction, fibre optimisation, protein adequacy, and omega-3 for triglyceride reduction. The oestrobolome concept has strong mechanistic plausibility but is graded moderate because the clinical trial data for fibre-mediated oestrogen modulation specifically are observational. Omega-3 for vasomotor symptoms and flaxseed for oestrogen modulation remain in the emerging category.
Outcomes data
- Alcohol reduction: Chen et al. (2011) demonstrated that each 10 g/day reduction in alcohol intake is associated with approximately 10 per cent reduction in breast cancer risk. Clinical experience suggests sleep quality improvements may be observed within the first few weeks.
- Fibre: meeting the SACN 30 g/day target is associated with significant reductions in cardiovascular disease, type 2 diabetes, and colorectal cancer risk (SACN, 2015). Emerging evidence suggests oestrobolome diversity improvements may become apparent within several weeks of sustained dietary change.
- Protein: Nunes et al. (2022) confirmed that intakes above 1.2 g/kg/day, combined with resistance exercise, consistently improve lean body mass.
- Omega-3: Chen et al. (2023) demonstrated significant triglyceride reductions with omega-3 supplementation in postmenopausal women. Anti-inflammatory effects are generally expected after approximately 8-12 weeks at 1-2 g/day (Calder, 2017).
When to seek professional support
NICE NG23 provides the clinical framework for identifying when patients require medical referral beyond nutritional support. Practitioners should refer promptly to a GP or menopause specialist when:
- Symptoms are severe and disabling despite lifestyle and dietary optimisation, particularly if the patient is considering or may benefit from HRT.
- The patient is under 45 with menopausal symptoms, as this may indicate premature ovarian insufficiency requiring specialist assessment and early HRT consideration for bone and cardiovascular protection.
- The patient is under 40 with menopausal symptoms -- this requires urgent specialist referral.
- Abnormal uterine bleeding occurs (postmenopausal bleeding, intermenstrual bleeding, or sudden change in bleeding pattern) -- this requires urgent gynaecological assessment to exclude endometrial pathology.
- Mental health symptoms are severe, including persistent low mood, suicidal ideation, or self-harm. In a crisis, direct the patient to call 999 (emergency), NHS 111 (urgent non-emergency), Samaritans on 116 123 (free, 24/7), or text SHOUT to 85258.
- Cardiovascular risk factors are present (hypertension, dyslipidaemia, family history of premature cardiovascular disease, diabetes) -- these require medical assessment and management alongside nutritional support. For guidance on cardiometabolic and hormonal lab panels, the lab-ordering framework covers test selection and sequencing.
- Osteoporosis risk is elevated (family history, low BMI, prolonged amenorrhoea, corticosteroid use) -- DEXA scanning and pharmacological intervention may be indicated.
Functional medicine nutrition is a complement to, not a replacement for, evidence-based medical care. The dietary interventions described in this article work synergistically with HRT where indicated, and do not contraindicate any standard medical treatment.
A four-pillar framework for perimenopausal nutrition
Perimenopause is not a disease. It is a physiological transition. But it is a transition whose symptoms and long-term health consequences are profoundly influenced by what a woman eats, drinks, and does with her body during these years.
The four dietary pillars -- reducing alcohol, optimising fibre, increasing protein, and ensuring adequate omega-3 intake -- are not exotic interventions. They are basic nutritional medicine, grounded in physiology and supported by systematic reviews. Yet they remain underutilised in clinical practice, overshadowed by a false dichotomy between "just get on with it" and "here is a prescription."
For Sarah, the protocol was straightforward. She reduced her wine from fourteen units to four per week. She added ground flaxseed to porridge and an extra portion of vegetables daily, raising fibre from 16 to 32 grams. She restructured meals around protein -- eggs at breakfast, sardines at lunch, legumes and fish at dinner -- reaching 90 grams per day. She began eating mackerel twice weekly and added an EPA-predominant omega-3 supplement on non-fish days.
Within six weeks, her night sweats had reduced from four nights a week to one. Her fasting glucose dropped to 5.2 mmol/L. She lost two inches from her waist circumference without changing her exercise routine. Her mood, she said, was "recognisable again." That is what happens when you address the underlying physiology rather than only treating the symptoms.
Practitioner summary
- Quantify alcohol intake honestly and communicate the dose-response clearly: each additional 10 g/day of ethanol increases breast cancer risk by approximately 10%. Target fewer than 7 units per week with at least three alcohol-free days.
- Frame alcohol reduction as a positive oestrogen-management strategy -- it reduces hepatic clearance burden and shifts metabolite ratios away from genotoxic 4-hydroxyoestrone.
- Increase dietary fibre to the SACN target of 30 g/day, building gradually over 2-4 weeks. Prioritise diversity (30 different plant foods per week) and include one cruciferous vegetable daily for sulforaphane-mediated oestrogen detoxification support.
- Add 2 tablespoons of ground flaxseed daily for lignans and soluble fibre to modulate oestrobolome activity.
- Raise protein to a minimum of 1.2 g/kg/day (up to 1.6 g/kg/day for active women), distributed across at least three meals with 25-30 g per meal to cross the leucine threshold for muscle protein synthesis.
- Combine protein targets with progressive resistance training at least twice weekly -- protein without mechanical loading is insufficient to reverse oestrogen-driven sarcopenia.
- Target 1-2 g combined EPA+DHA daily (two portions of oily fish per week, or algal supplement). Prioritise EPA-predominant formulations for anti-inflammatory and mood benefits.
- Ensure adequate vitamin D status (test 25[OH]D, apply BMI-adjusted dosing where indicated) as vitamin D and omega-3 have synergistic anti-inflammatory effects.
- Refer promptly to a GP or menopause specialist for symptoms under age 45 (premature ovarian insufficiency), abnormal uterine bleeding, or severe symptoms unresponsive to dietary optimisation.
- These dietary levers complement HRT and do not contraindicate any standard medical treatment -- communicate this clearly to patients already on or considering pharmacological management.
References
- [1]Baker, J.M., Al-Nakkash, L (2017) ‘and Herbst-Kralovetz, M.M’, (. doi:10.1016/j.maturitas.2017.06.025
- [2]Bauer, J., Biolo, G., Cederholm, T., Cesari, M., Cruz-Jentoft, A.J., Morley, J.E., Phillips, S., Sieber, C., Stehle, P., Teta, D., Visvanathan, R., Volpi, E (2013) ‘and Boirie, Y’, (. doi:10.1016/j.jamda.2013.05.021
- [3]Calder, P.C (2017) ‘(2017) 'Omega-3 fatty acids and inflammatory processes: from molecules to man', Biochemical Society Transactions, 45(5), pp’, 1105-1115. doi:10.1042/BST20160474
- [4]Chen, C., Gong, X., Yang, X., Shang, X., Du, Q., Liao, Q., Xie, R., Chen, Y (2023) ‘and Xu, J’, (. doi:10.1016/j.clinthera.2022.11.007
- [5]Chen, W.Y., Rosner, B., Hankinson, S.E., Colditz, G.A (2011) ‘and Willett, W.C’, (. doi:10.1001/jama.2011.1590
- [6]Erdelyi, A., Palfi, E., Tuu, L., Nas, K., Szucs, Z., Torok, M., Jakab, A (2023) ‘and Varbiro, S’, (. doi:10.3390/nu16010027
- [7]Harlow, S.D., Gass, M., Hall, J.E., Lobo, R., Maki, P., Rebar, R.W., Sherman, S., Sluss, P.M (2012) ‘and de Villiers, T.J’, (. doi:10.1097/gme.0b013e31824d8f40
- [8]Iqbal, A.Z., Wu, S.K., Zailani, H., Chiu, W.C., Liu, W.C., Su, K.P (2023) ‘and Lee, S.D’, (. doi:10.3390/nu15194231
- [9]National Institute for Health and Care Excellence (2015; updated 2024) *Menopause: identification and management* (2015) ‘NICE guideline NG23’, Available at: https://www.nice.org.uk/guidance/ng23 (Accessed: 25 May. www.nice.org.uk
- [10]Nunes, E.A., Colenso-Semple, L., McKellar, S.R., Yau, T., Courtney-Martin, G., Szeto, H.H., Phillips, S.M (2022) ‘and McMaster University (2022) 'Systematic review and meta-analysis of protein intake to support muscle mass and function in healthy adults', Journal of Cachexia, Sarcopenia and Muscle, 13(2), pp’, 795-810. doi:10.1002/jcsm.12922
- [11]Playdon, M.C., Coburn, S.B., Moore, S.C., Brinton, L.A., Wentzensen, N., Anderson, G., Wallace, R., Falk, R.T., Pfeiffer, R., Xu, X (2018) ‘and Trabert, B’, (. doi:10.1038/bjc.2017.419
- [12]Plottel, C.S (2011) ‘and Blaser, M.J’, (. doi:10.1016/j.chom.2011.10.003
- [13]Santoro, N., Roeca, C., Peters, B.A (2021) ‘and Neal-Perry, G’, (. doi:10.1210/clinem/dgaa764
- [14]Scientific Advisory Committee on Nutrition (2015) *Carbohydrates and Health* (2015) ‘London: The Stationery Office’, Available at: https://www.gov.uk/government/publications/sacn-carbohydrates-and-health-report (Accessed: 25 May. www.gov.uk
- [15]Sohi, I., Rehm, J., Saab, M., Virmani, L., Franklin, A., Sanchez, G., Shield, K (2024) ‘et al’, (. doi:10.1111/acer.15493
Written by

Chris Massamba
Functional Nutritionist & Health Coach · Dip CNM · FMCHC
Medical disclaimer: The content in this article is for informational purposes only and does not constitute medical advice, diagnosis, or treatment. Always consult a qualified healthcare professional before making any changes to your health regimen. Individual results may vary. If you are experiencing a medical emergency, please contact 999 immediately.
Related Protocols
- Hormonal Balancemoderate
- Thyroid Supportmoderate
- Adrenal Supportmoderate